
文献分享-两种芥菜杂交形成稳定的多倍体
异源四倍体芥菜(AABB)和埃塞俄比亚芥(BBCC)杂交后形成稳定的杂交种(BBAC)
Introduction
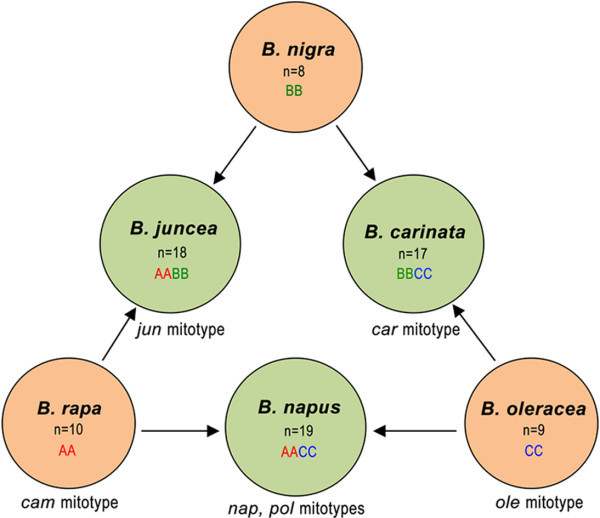
动物植物中都有多倍体的存在,开花植物中更为常见。二倍体杂交形成的异源四倍体为初级多倍体,两个异源四倍体杂交产生次级多倍体。祖先关系表明:AC亚基因组关系更近,B亚基因组相对较远6Ma。尽管B基因组物种从A/C谱系中分离出来约6 Ma,但古多倍体基因组的比较揭示了基因内容和序列同一性的广泛保守性。
- 在山羊草属研究中发现,有共享基因组的异源四倍体更容易杂交。两个异源四倍体杂交初期核型不稳定,共享的基因组能为次级异源四倍体形成提供缓冲。
- 伴随多倍体的基因组变化的分子分析使我们在理解初级多倍体如何形成新的、稳定的基因组方面的重大突破。然而,次级多倍体如何形成稳定的重组基因组是未知的。阐明这一过程将加深我们对十字花科内微进化分化的理解,并有助于系统发育的重建。
研究对象
芸薹属植物是研究种间杂交和多倍体的重要模式植物。作者选取了芥菜(AABB)和埃塞俄比亚芥(BBCC)作为亲本,自交多代,统计结实率、花粉活力和构建核型。
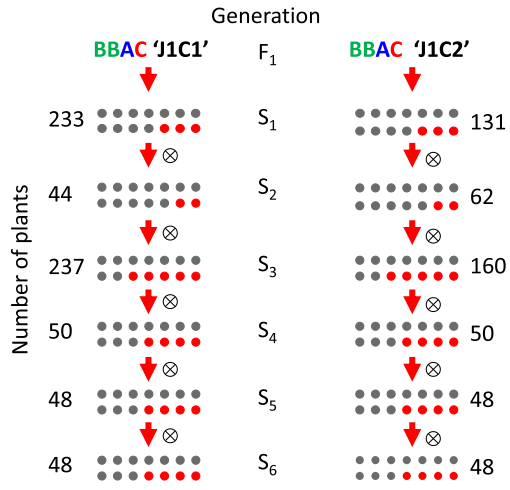
科学问题:确定BBAC杂交后代在多代中的基因组稳定性和生育能力,以了解是否可以在后代中恢复稳定、产生可育的杂交后代,如果可以,通过何种机制恢复。
自交选育
亲本:将芥菜与与两种埃塞俄比亚芥杂交产生,J1C1(20)和J1C2(24)两种杂交后代。
环境:条件不同的温室和大田里生长。
后代亲本选择:选择可育性最强的2-5个植株作为下一代亲本。
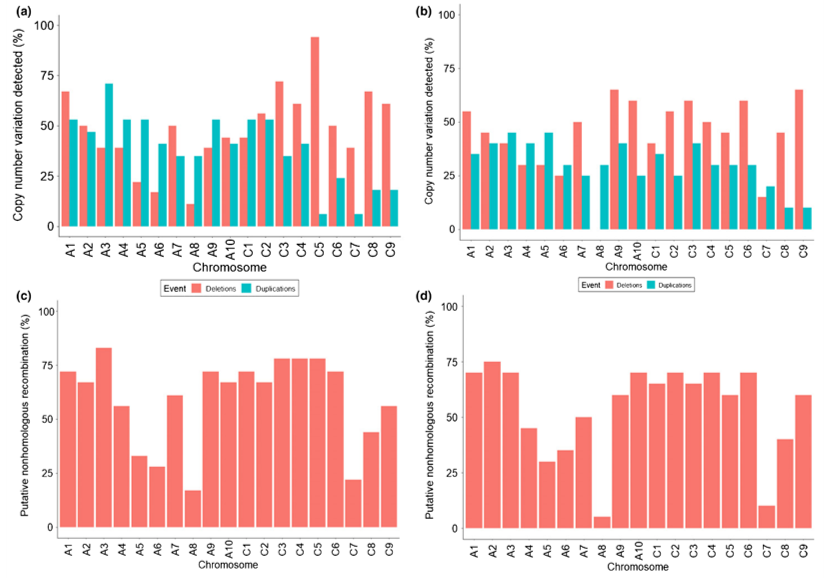
拷贝数变化和非同源重组
S1代中发生了大量的非同源重组。
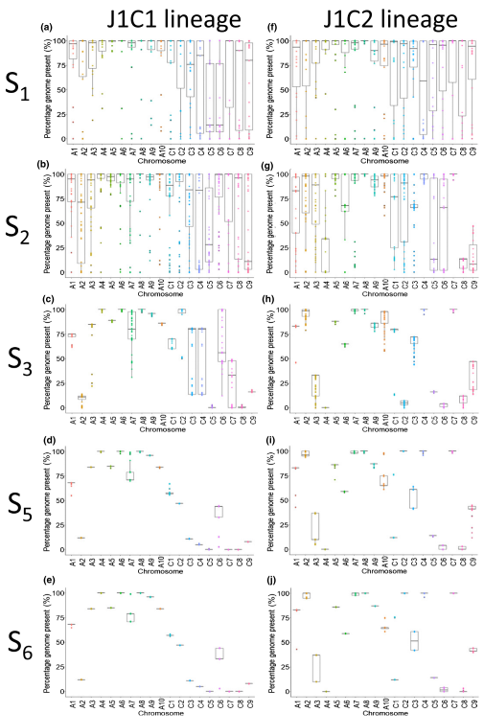
后代中基因组的保留比例
与S2代相比,S3代中不同染色体存在的基因组百分比变异减少,与“杂合”染色体重排固定增加的预期一致。
在后代可育性的统计中,A和C基因组中任何大规模(>0.5 Mbp)基因组重排对育性没有显著影响。 但J1C1中丢失A3染色体, J1C2中丢失A1染色体会减小可育性。
花粉活力和染色体数目
到了S4代杂种的花粉活力和种子育性都有所提高。两种基因型间种子育性有差异。
两种基因型植株的染色体数目均在32~36条之间,平均为34条。
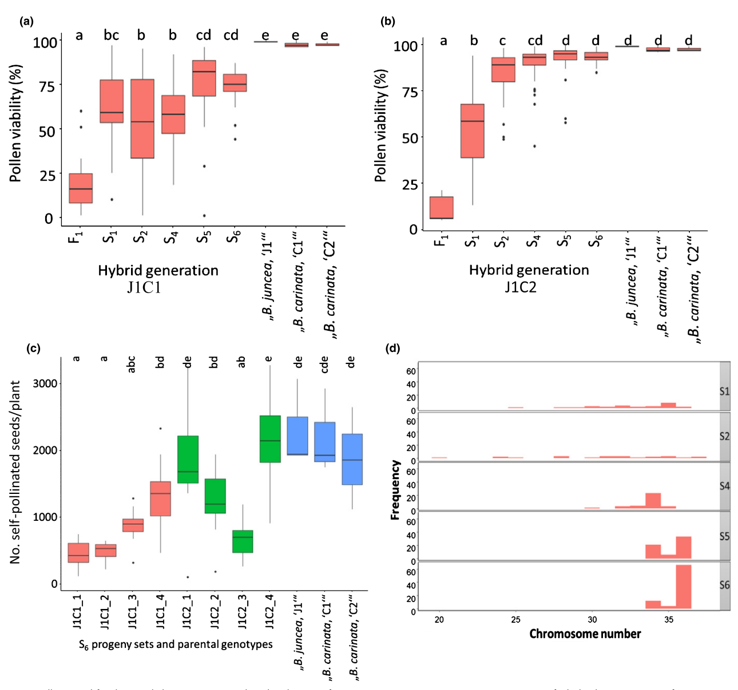
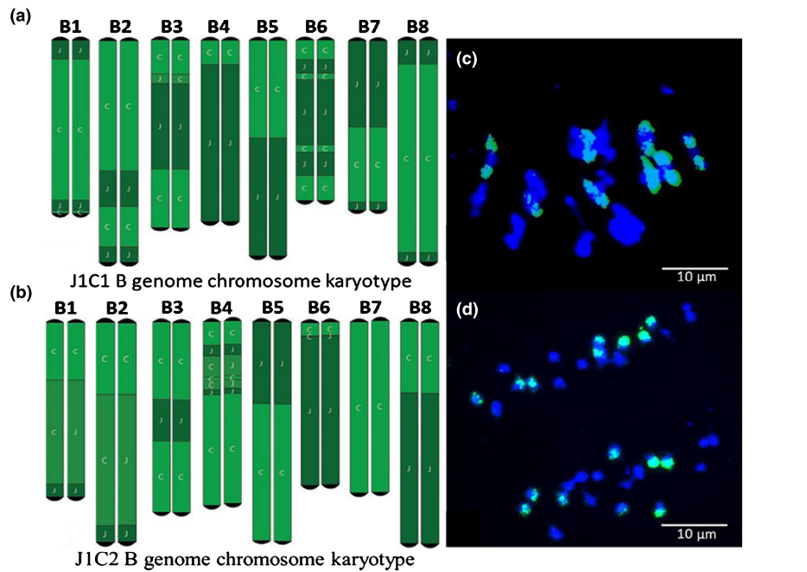
B基因组未与A/C基因组重组
在S5和S6代中,所有16条B基因组染色体均存在,且未与任何A或C基因组染色体重组。通过对有丝分裂和减数分裂染色体制备物进行基因组原位杂交,在中期I始终观察到16条B染色体正确配对产生的预期8个二价体,并且这些染色体在后期I正确分离(8条染色体位于对极)(d图)。
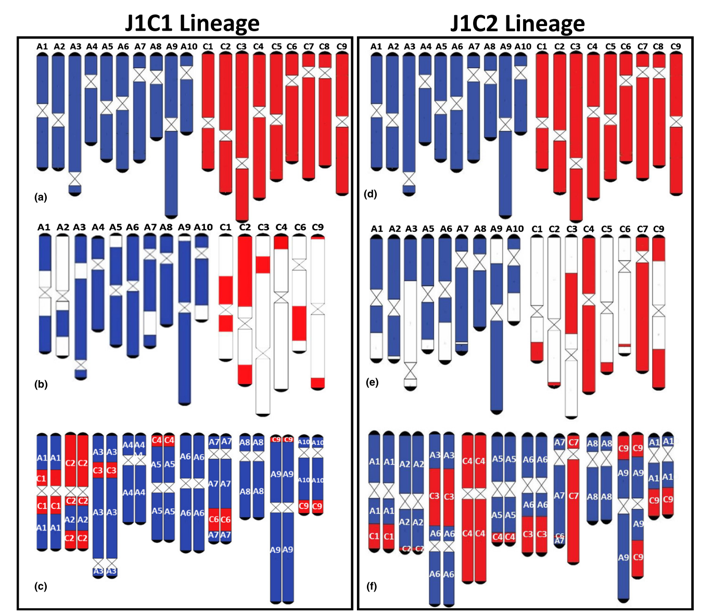
AC基因组高度重组
A和C基因组之间高度重组,甚至包括染色体的
增加和丢失。
这种重组没有明确的选择压力。
在两个谱系中,A基因组的保留都有明显的偏向。
在每个谱系中,每个部分同源染色体区域在
最终A和C基因组核型中以恰好一个拷贝存在,不存在都保留和都缺失的情况。
结论
1. 通过自花授粉和育性选择可以产生稳定的、可育的、具有新核型的杂种。
2. 在BBAC杂种中,A和C基因组之间发生重组和重构,而B基因组保持不变,这些A/C重排似乎在S5/6代被固定,伴随着育性和减数分裂稳定性的恢复,产生“真正育种”后代
自我感悟
1、染色体数量或者核型在植物居群中是不稳定的。
2、AABB和BBCC异源四倍体杂交后,后代的染色体数据是逐渐稳定的,可育性是逐渐增加的。
3、染色体数目的减少是降解还是融合,增加是断裂。
4、可育性是因为选择,每代选择可育的个体的种子。
5、A亚基因组和C亚基因组的重排形成的最终核型中,每一个同源染色体区域都精确的保留了一个拷贝。
6、大多是交换都发生在同源度非常高的区域。
7、A和C大部分重组发生在S1S2减数分裂中,并在后代中随机固定。
8、在我们的研究中,我们观察到BBAC杂种在自花授粉六代后,B基因组和A基因组之间以及B基因组和C基因组之间完全缺乏重组,是因为AC之间形成了强力的配对。
9、之前小麦的研究中异源四倍体RRAB直到16代都没有形成稳定的核型,可能因为小麦中的Ph1基因能够完全阻止同源重组,相比之下,芸薹属没有强有力的阻止AC之间部同源重组。
10、BBAC谱系中,C基因组更偏向缺失。
名词解释
同源性指的是由于物种形成而从同一起源分化出来的染色体或染色体片段,通过异源多倍体,即异源多倍体被带回到同一基因组中。
同源重组:父母本的同源染色体重组。例如人类来自父亲的1号和自母亲的1号染色体重组。
非同源重组:非同源染色体之间的重组,例如人类的1号和2号染色体重组。